Изучением строения организмов, а также растений животных и человека занимается раздел биологии, называемый цитологией. Ученые установили, что содержимое клетки, которое находится внутри нее, построено довольно сложно. Его окружает так называемый поверхностный аппарат, в состав которого входят наружная клеточная мембрана, надмембранные структуры: гликокаликс и а также микронити, пеликула и микротрубочки, образующие её подмембранный комплекс.
В данной статье мы изучим строение и функции наружной клеточной мембраны, входящей в поверхностный аппарат различных видов клеток.
Какие функции выполняет наружная клеточная мембрана
Как было описано ранее, наружная мембрана является частью поверхностного аппарата каждой клетки, который успешно отделяет ее внутреннее содержимое и защищает клеточные органеллы от неблагоприятных условий внешней среды. Еще одна функция - это обеспечение обмена веществ между клеточным содержимым и тканевой жидкостью, поэтому наружная клеточная мембрана осуществляет транспорт молекул и ионов, поступающих в цитоплазму, а также помогает удалять шлаки и избыток токсичных веществ из клетки.

Строение клеточной мембраны
Мембраны, или плазмалеммы различных типов клеток сильно отличаются между собой. Главным образом, химическим строением, а также относительным содержанием в них липидов, гликопротеидов, белков и, соответственно, характером рецепторов, находящихся в них. Наружная которой определяются прежде всего индивидуальным составом гликопротеидов, берет участие в распознавании раздражителей внешней среды и в реакциях самой клетки на их действия. С белками и гликолипидами клеточных мембран могут взаимодействовать некоторые виды вирусов, вследствие чего они проникают в клетку. Вирусы герпеса и гриппа могут использовать для построения свой защитной оболочки.
А вирусы и бактерии, так называемые бактериофаги, прикрепляются к мембране клетки и в месте контакта растворяют ее с помощью особого фермента. Затем в образовавшееся отверстие проходит молекула вирусной ДНК.
Особенности строения плазмалеммы эукариот
Напомним, что наружная клеточная мембрана выполняет функцию транспорта, то есть переноса веществ в и из нее во внешнюю среду. Для осуществления такого процесса необходимо специальное строение. Действительно, плазмалемма представляет собой постоянную, универсальную для всех систему поверхностного аппарата. Это тоненькая (2-10 Нм), но достаточно плотная многослойная пленка, которая покрывает всю клетку. Её строение было изучено в 1972 году такими учеными, как Д. Сингер и Г. Николсон, ими же создана жидкостно-мозаичная модель клеточной мембраны.
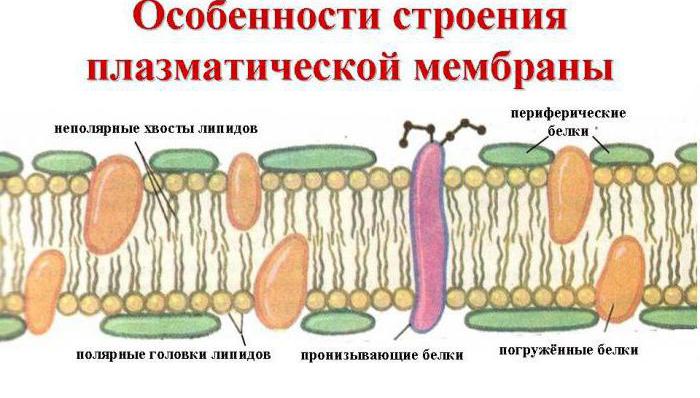
Главные химические соединения, которые её образуют - это упорядоченно расположенные молекулы белков и определенных фосфолипидов, которые вкраплены в жидковатую липидную среду и напоминают мозаику. Таким образом, клеточная мембрана состоит из двух слоев липидов, неполярные гидрофобные «хвосты» которых находятся внутри мембраны, а полярные гидрофильные головки обращены к цитоплазме клетки и к межклеточной жидкости.
Слой липидов пронизывается крупными белковыми молекулами, образующими гидрофильные поры. Именно через них транспортируются водные растворы глюкозы и минеральных солей. Некоторые белковые молекулы находятся как на внешней, так и на внутренней поверхности плазмалеммы. Таким образом, на наружной клеточной мембране в клетках всех организмов, имеющих ядра, находятся молекулы углеводов, связанные ковалентными связями с гликолипидами и гликопротеидами. Содержание углеводов в клеточных мембранах колеблется от 2 до 10%.

Строение плазмалеммы прокариотических организмов
Наружная клеточная мембрана у прокариот выполняет сходные функции с плазмалеммами клеток ядерных организмов, а именно: восприятие и передача информации, поступающей из внешней среды, транспорт ионов и растворов в клетку и из нее, защита цитоплазмы от чужеродных реагентов извне. Она может образовывать мезосомы - структуры, возникающие при впячивании плазмалеммы внутрь клетки. На них могут находиться ферменты, участвующие в метаболических реакциях прокариот, например, в репликации ДНК, синтезе белков.
Мезосомы также содержат окислительно-восстановительные ферменты, а у фотосинтетиков находятся бактериохлорофилл (у бактерий) и фикобилин (у цианобактерий).
Роль наружных мембран в межклеточных контактах
Продолжая отвечать на вопрос, какие функции выполняет наружная клеточная мембрана, остановимся на ее роли в У растительных клеток в стенках наружной клеточной мембраны образуются поры, переходящие в целлюлозный слой. Через них возможен выход цитоплазмы клетки наружу, такие тонкие каналы называют плазмодесмами.

Благодаря им связь между соседними растительными клетками очень прочная. У клеток человека и животных места контактов соседних клеточных мембран называются десмосомами. Они характерны для эндотелиальных и эпителиальных клеток, а также встречаются у кардиомиоцитов.
Вспомогательные образования плазмалеммы
Разобраться, чем отличаются растительные клетки от животных, помогает изучение особенностей строения их плазмалемм, которые зависят от того, какие функции выполняет наружная клеточная мембрана. Над ней у животных клеток находится слой гликокаликс. Он образован молекулами полисахаридов, связанных с белками и липидами наружной клеточной мембраны. Благодаря гликокаликсу между клетками возникает адгезия (слипание), приводящая к образованию тканей, поэтому он принимает участие в сигнальной функции плазмалеммы - распознавании раздражителей внешней среды.
Как осуществляется пассивный транспорт определенных веществ через клеточные мембраны
Как было уже сказано ранее, наружная клеточная мембрана участвует в процессе транспортировки веществ между клеткой и внешней средой. Существует два вида переноса через плазмалемму: пассивный (дифузионный) и активный транспорт. К первому относится диффузия, облегченная диффузия и осмос. Движение веществ по градиенту концентрации зависит, прежде всего, от массы и величины молекул, проходящих через клеточную мембрану. Например, мелкие неполярные молекулы легко растворяются в среднем липидном слое плазмалеммы, продвигаются через нее и оказываются в цитоплазме.

Крупные молекулы органических веществ проникают в цитоплазму с помощью специальных белков-переносчиков. Они имеют видовую специфичность и, соединяясь с частицей или ионом, без затрат энергии пассивно переносят их через мембрану по градиенту концентрации (пассивный транспорт). Этот процесс лежит в основе такого свойства плазмалеммы, как избирательная проницаемость. В процессе энергия молекул АТФ не используется, и клетка сберегает её на другие метаболические реакции.
Активный транспорт химических соединений через плазмалемму
Так как наружная клеточная мембрана обеспечивает перенос молекул и ионов из внешней среды внутрь клетки и обратно, становится возможным вывод продуктов диссимиляции, являющихся токсинами, наружу, то есть в межклеточную жидкость. происходит против градиента концентрации и требует использования энергии в виде молекул АТФ. В нем также участвуют белки-переносчики, называемые АТФ-азами, являющиеся одновременно и ферментами.

Примером такого транспорта служит натрий-калиевый насос (ионы натрия переходят из цитоплазмы во внешнюю среду, а ионы калия закачиваются в цитоплазму). К нему способны эпителиальные клетки кишечника и почек. Разновидностями такого способа переноса служат процессы пиноцитоза и фагоцитоза. Таким образом, изучив, какие функции выполняет наружная клеточная мембрана, можно установить, что к процессам пино- и фагоцитоза способны гетеротрофные протисты, а также клетки высших животных организмов, например, лейкоциты.
Биоэлектрические процессы в клеточных мембранах
Установлено, что существует разность потенциалов между наружной поверхностью плазмалеммы (она заряжена положительно) и пристеночным слоем цитоплазмы, заряженным отрицательно. Ее назвали потенциалом покоя, и она присуща всем живым клеткам. А нервная ткань имеет не только потенциал покоя, но и способна к проведению слабых биотоков, которое называют процессом возбуждения. Наружные мембраны нервных клеток-нейронов, принимая раздражение от рецепторов, начинают менять заряды: ионы натрия массированно поступают внутрь клетки и поверхность плазмалеммы становится электроотрицательной. А пристеночный слой цитоплазмы вследствие избытка катионов получает положительный заряд. Это объясняет, по какой причине происходит перезарядка наружной клеточной мембраны нейрона, что вызывает проведение нервных импульсов, лежащих в основе процесса возбуждения.
Среди основных функций клеточной мембраны можно выделить барьерную, транспортную, ферментативную и рецепторную . Клеточная (биологическая) мембрана (она же плазмалемма, плазматическая или цитоплазматическая мембрана) ограждает содержимое клетки или ее органоидов от окружающей среды, обеспечивает избирательную проницаемость для веществ, на ней располагаются ферменты, а также молекулы, способные «улавливать» различные химические и физические сигналы.
Такая функциональность обеспечивается особым строением клеточной мембраны .
В эволюции жизни на Земле клетка вообще могла образоваться лишь после появления мембраны, которая отделила и стабилизировала внутреннее содержимое, не дало ему распасться.
В плане поддержания гомеостаза (саморегуляции относительного постоянства внутренней среды) барьерная функция клеточной мембраны тесно связана с транспортной .
Малые молекулы способны проходить сквозь плазмалемму без всяких «помощников», по градиенту концентрации, т. е. из области с высокой концентрацией данного вещества в область с низкой концентрацией. Так, например, обстоит дело для газов, участвующих в дыхании. Кислород и углекислый газ диффундируют через клеточную мембрану в том направлении, где их концентрация в данный момент меньше.
Поскольку мембрана в основной своей части гидрофобна (из-за двойного липидного слоя), то полярные (гидрофильные) молекулы, даже малых размеров, зачастую не могут сквозь нее проникнуть. Поэтому ряд мембранных белков выполняет функцию переносчиков таких молекул, связываясь с ними и перенося через плазмалемму.
Интегральные (пронизывающие мембрану насквозь) белки часто работают по принципу открывающихся и закрывающихся каналов. Когда какая-либо молекула подходит к такому белку, то он соединяется с ней, и канал открывается. Это вещество или другое проходит через белковый канал, после чего его конформация меняется, и канал закрывается для этого вещества, но может открыться для пропускания другого. По такому принципу работает натрий-калиевый насос, закачивающий в клетку ионы калия и выкачивающий из нее ионы натрия.
Ферментативная функция клеточной мембраны в большей степени реализована на мембранах органоидов клетки. Большинство синтезируемых в клетке белков выполняют ферментативную функцию. «Усаживаясь» на мембрану в определенном порядке, они организуют конвейер, когда продукт реакции, катализируемый одним белком-ферментом, переходит к следующему. Такой «конвейер» стабилизируют поверхностные белки плазмалеммы.
Несмотря на универсальность строения всех биологических мембран (построены по единому принципу, почти одинаковы у всех организмов и у разных мембранных клеточных структур), их химический состав все же может отличаться. Бывают более жидкие и более твердые, на одних больше определенных белков, на других меньше. Кроме того, отличаются и разные стороны (внутренняя и наружная) одной и той же мембраны.
У мембраны, которая окружает клетку (цитоплазматической) на внешней стороне располагается множество углеводных цепей, прикрепленных к липидам или белкам (в результате образуются гликолипиды и гликопротеины). Многие из таких углеводов выполняют рецепторную функцию , будучи восприимчивыми к определенным гормонам, улавливая изменения физических и химических показателей в окружающей среде.
Если, например, гормон соединяется со своим клеточным рецептором, то углеводная часть молекулы-рецептора изменяет свое строение, вслед за ней изменяет строение и связанная с ней белковая часть, пронизывающая мембрану. На следующем этапе в клетке запускаются или приостанавливаются различные биохимические реакции, т. е. меняется ее метаболизм, начинается клеточный ответ на «раздражитель».
Кроме перечисленных четырех функций клеточной мембраны выделяют и другие: матричную, энергетическую, маркировачную, формирование межклеточных контактов и др. Однако их можно рассмотреть как «подфункции» уже рассмотренных.
Клеточная мембрана также называется плазматической (или цитоплазматической) мембраной и плазмалеммой. Данная структура не только отделяет внутреннее содержимое клетки от внешней среды, но также входит с состав большинства клеточных органелл и ядра, в свою очередь отделяя их от гиалоплазмы (цитозоля) - вязко-жидкой части цитоплазмы. Договоримся называть цитоплазматической мембраной ту, которая отделяет содержимое клетки от внешней среды. Остальными терминами обозначать все мембраны.
В основе строения клеточной (биологической) мембраны лежит двойной слой липидов (жиров). Формирование такого слоя связано с особенностями их молекул. Липиды не растворяются в воде, а по-своему в ней конденсируются. Одна часть отдельно взятой молекулы липида представляет собой полярную головку (она притягивается водой, т. е. гидрофильна), а другая - пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.
В результате образуется двойной липидный слой, в котором неполярные хвосты находятся внутри (обращены друг к другу), а полярные головки обращены наружу (к внешней среде и цитоплазме). Поверхность такой мембраны гидрофильна, а внутри она гидрофобна.
В клеточных мембранах среди липидов преобладают фосфолипиды (относятся к сложным липидам). Их головки содержат остаток фосфорной кислоты. Кроме фосфолипидов есть гликолипиды (липиды + углеводы) и холестерол (относится к стеролам). Последний придает мембране жесткость, размещаясь в ее толще между хвостами остальных липидов (холестерол полностью гидрофобный).
За счет электростатического взаимодействия, к заряженным головкам липидов присоединяются некоторые молекулы белков, которые становятся поверхностными мембранными белками. Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Таким образом, клеточная мембрана состоит из двойного слоя липидов, поверхностных (периферических), погруженных (полуинтегральных) и пронизывающих (интегральных) белков . Кроме того, некоторые белки и липиды с внешней стороны мембраны связаны с углеводными цепями.

Это жидкостно-мозаичная модель строения мембраны была выдвинута в 70-х годах XX века. До этого предполагалась бутербродная модель строения, согласно которой липидный бислой находится внутри, а с внутренней и наружной стороны мембрана покрыта сплошными слоями поверхностных белков. Однако накопление экспериментальных данных опровергло эту гипотезу.
Толщина мембран у разных клеток составляет около 8 нм. Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Разрывы клеточной мембраны легко сливаются из-за физико-химических особенностей липидного бислоя. В плоскости мембраны липиды и белки (если только они не закреплены цитоскелетом) перемещаются.
Функции клеточной мембраны
Большинство погруженных в клеточную мембрану белков выполняют ферментативную функцию (являются ферментами). Часто (особенно в мембранах органоидов клетки) ферменты располагаются в определенной последовательности так, что продукты реакции, катализируемые одним ферментом, переходят ко второму, затем третьему и т. д. Образуется конвейер, который стабилизируют поверхностные белки, т. к. не дают ферментам плавать вдоль липидного бислоя.
Клеточная мембрана выполняет отграничивающую (барьерную) от окружающей среды и в то же время транспортную функции. Можно сказать, это ее самое главное назначение. Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность).
При этом транспорт веществ происходит различными способами. Транспорт по градиенту концентрации предполагает передвижение веществ из области с их большей концентрацией в область с меньшей (диффузия). Так, например, диффундируют газы (CO 2 , O 2).
Бывает также транспорт против градиента концентрации, но с затратой энергии.
Транспорт бывает пассивным и облегченным (когда ему помогает какой-нибудь переносчик). Пассивная диффузия через клеточную мембрану возможна для жирорастворимых веществ.
Есть особые белки, делающие мембраны проницаемыми для сахаров и других водорастворимых веществ. Такие переносчики соединяются с транспортируемыми молекулами и протаскивают их через мембрану. Так переносится глюкоза внутрь эритроцитов.
Пронизывающие белки, объединяясь, могут образовывать пору для перемещения некоторых веществ через мембрану. Такие переносчики не перемещаются, а образуют в мембране канал и работают аналогично ферментам, связывая определенное вещество. Перенос осуществляется благодаря изменению конформации белка, благодаря чему в мембране образуются каналы. Пример - натрий-калиевый насос.
Транспортная функция клеточной мембраны эукариот также реализуется за счет эндоцитоза (и экзоцитоза). Благодаря этим механизмам в клетку (и из нее) попадают крупные молекулы биополимеров, даже целые клетки. Эндо- и экзоцитоз характерны не для всех клеток эукариот (у прокариот его вообще нет). Так эндоцитоз наблюдается у простейших и низших беспозвоночны; у млекопитающих лейкоциты и макрофаги поглощают вредные вещества и бактерии, т. е. эндоцитоз выполняет защитную функцию для организма.
Эндоцитоз делится на фагоцитоз (цитоплазма обволакивает крупные частицы) и пиноцитоз (захват капелек жидкости с растворенными в ней веществами). Механизм этих процессов приблизительно одинаков. Поглощаемые вещества на поверхности клеток окружаются мембраной. Образуется пузырек (фагоцитарный или пиноцитарный), который затем перемещается внутрь клетки.
Экзоцитоз - это выведение цитоплазматической мембраной веществ из клетки (гормонов, полисахаридов, белков, жиров и др.). Данные вещества заключаются в мембранные пузырьки, которые подходят к клеточной мембране. Обе мембраны сливаются и содержимое оказывается за пределами клетки.
Цитоплазматическая мембрана выполняет рецепторную функцию. Для этого на ее внешней стороне располагаются структуры, способные распознавать химический или физический раздражитель. Часть пронизывающих плазмалемму белков с наружней стороны соединены с полисахаридными цепочками (образуя гликопротеиды). Это своеобразные молекулярные рецепторы, улавливающие гормоны. Когда конкретный гормон связывается со своим рецептором, то изменяет его структуру. Это в свою очередь запускает механизм клеточного ответа. При этом могут открываться каналы, и в клетку могут начать поступать определенные вещества или выводиться из нее.
Рецепторная функция клеточных мембран хорошо изучена на основе действия гормона инсулина. При связывании инсулина с его рецептором-гликопротеидом происходит активация каталитической внутриклеточной части этого белка (фермента аденилатциклазы). Фермент синтезирует из АТФ циклическую АМФ. Уже она активирует или подавляет различные ферменты клеточного метаболизма.
Рецепторная функция цитоплазматической мембраны также включает распознавание соседних однотипных клеток. Такие клетки прикрепляются друг к другу различными межклеточными контактами.
В тканях с помощью межклеточных контактов клетки могут обмениваться между собой информацией с помощью специально синтезируемых низкомолекулярных веществ. Одним из примеров подобного взаимодействия является контактное торможение, когда клетки прекращают рост, получив информацию, что свободное пространство занято.
Межклеточные контакты бывают простыми (мембраны разных клеток прилегают друг к другу), замковыми (впячивания мембраны одной клетки в другую), десмосомы (когда мембраны соединены пучками поперечных волокон, проникающих в цитоплазму). Кроме того, есть вариант межклеточных контактов за счет медиаторов (посредников) - синапсы. В них сигнал передается не только химическим, но и электрическим способом. Синапсами передаются сигналы между нервными клетками, а также от нервных к мышечным.
text_fields
text_fields
arrow_upward
Клетки отделены от внутренней среды организма клеточной или плазматической мембраной.
Мембрана обеспечивает:
1) Избирательное проникновение в клетку и из нее молекул и ионов, необходимых для выполнения специфических функций клеток;
2) Избирательный транспорт ионов через мембрану, поддерживая трансмембранную разницу электрического потенциала;
3) Специфику межклеточных контактов.
Благодаря наличию в мембране многочисленных рецепторов, воспринимающих химические сигналы - гормоны, медиаторы и другие биологически активные вещества, она способна изменять метаболическую активность клетки. Мембраны обеспечивают специфику иммунных проявлений, благодаря наличию на них антигенов - структур, вызывающих образование антител, способных специфически связываться с этими антигенами.
Ядро и органеллы клетки также отделены от цитоплазмы мембранами, которые предупреждают свободное движение воды и растворенных в ней веществ из цитоплазмы в них и наоборот. Это создает условия для разделения биохимических процессов, протекающих в различных отсеках (компартментах) внутри клетки.
Структура мембраны клетки
text_fields
text_fields
arrow_upward
Мембрана клетки - эластичная структура, толщиной от 7 до 11 нм (рис.1.1). Она состоит, в основном, из липидоа и белков. От 40 до 90% всех липидов составляют фосфолипиды — фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, сфингомиелин и фосфатидилинозит. Важным компонентом мембраны являются гликолипиды, представленные цереброзидами, сульфатидами, ганглиозидами и холестерином.
Рис. 1.1 Организация мембраны.
Основной структурой мембраны клетки является двойной слой фосфолипидных молекул. За счет гидрофобных взаимодействий углеводные цепочки липидных молекул удерживаются друг возле друга в вытянутом состоянии. Группы же фосфолипидных молекул обоих слоев взаимо действуют с белковыми молекулами, погруженными в липидную мембрану. Благодаря тому, что большинство липидных компонентов бислоя находится в жидком состоянии, мембрана обладает подвижностью, совершает волнообразные движения. Ее участки, а также белки, погруженные в липидный бислой, перемешаются из одной ее части в другую. Подвижность (текучесть) мембран клеток облегчает процессы транспорта веществ через мембрану.
Белки мембраны клеток представлены, в основном, гликопротеинами. Различают:
интегральные белки
, проникающие через всю толщу мембраны и
периферические белки
, прикрепленные только к поверхности мембраны, в основном, к внутренней ее части.
Периферические белки почти все функционируют как энзимы (ацетилхолинестераза, кислая и шелочная фосфатазы и др.). Но некоторые энзимы также представлены интегральными белками - АТФ-аза.
Интегральные белки обеспечивают селективный обмен ионов через каналы мембран между экстрацеллюлярной и интрацеллюлярной жидкостью, а также действуют как белки - переносчики крупных молекул.
Рецепторы и антигены мембраны могут быть представлены как интегральными, так и периферическими белками.
Белки, примыкающие к мембране с цитоплазматической стороны, относятся к цитоскелету клетки . Они могут прикрепляться к мембранным белкам.
Так, белок полосы 3 (номер полосы при электрофорезе белков) эритроцитарных мембран объединяется в ансамбль с другими молекулами цитоскелета - спектрином через низкомолекулярный белок анкирин (рис. 1.2).
Рис. 1.2 Схема расположения белков в примембранном цитоскелете эритроцитов.1 - спектрин; 2 - анкирин; 3 - белок полосы 3; 4 - белок полосы 4,1; 5 - белок полосы 4,9; 6 - олигомер актина; 7 - белок 6; 8 - гпикофорин А; 9 - мембрана.
Спектрин является основным белком цитоскелета, составляющим двумерную сеть, к которой прикрепляется актин.
Актин образует микрофиламенты, представляющие собой сократительный аппарат цитоскелета.
Цитоскелет позволяет клетке проявлять гибкоэластические свойства, обеспечивает дополнительную прочность мембраны.
Большинство интегральных белков - гликопротеины . Их углеводная часть выступает из клеточной мембраны наружу. Многие гликопротеины обладают большим отрицательным зарядом из-за значительного содержания сиаловой кислоты (например, молекула гликофорина). Это обеспечивает поверхности большинства клеток отрицательный заряд, способствуя отталкиванию других отрицательно заряженных объектов. Углеводные выступы гликопротеинов являются носителями антигенов групп крови, других антигенных детерминант клетки, они действуют как рецепторы, связывающие гормоны. Гликопротеины образуют адгезивные молекулы, обуславливающие прикрепление клеток одна к другой, т.е. тесные межклеточные контакты.
Особенности обмена веществ в мембране
text_fields
text_fields
arrow_upward
Мембранные компоненты подвержены многим метаболическим превращениям под влиянием ферментов, расположенных на их мембране или внутри ее. К ним относятся окислительные ферменты, играющие важную роль в модификации гидрофобных элементов мембран - холестерина и др. В мембранах же при активации ферментов - фосфолипаз происходит образование из арахидоновой кислоты биологически активных соединений - простагландинов и их производных. В результате активации метаболизма фосфолипидов в мембране образуются тромбоксаны, лейкотриены, оказывающие мощное воздействие на адгезию тромбоцитов, процесс воспаления и др.
В мембране непрерывно протекают процессы обновления ее компонентов . Так, время жизни мембранных белков колеблется от 2 до 5 дней. Однако в клетке существуют механизмы, обеспечивающие доставку вновь синтезированных молекул белка к мембранным рецепторам, облегчающим встраивание белка в мембрану. «Узнавание» данного рецептора вновь синтезированным белком облегчается образованием сигнального пептида, помогающего найти на мембране рецептор.
Липиды мембраны отличаются также значительной скоростью обмена
, что требует для синтеза этих компонентов мембраны большого количества жирных кислот.
На специфику липидного состава мембран клеток влияют изменения среды обитания человека, характера его питания.
Например, увеличение в пище жирных кислот с ненасыщенными связями увеличивает жидкое состояние липидов мембран клеток различных тканей, приводит к благоприятному для функции мембраны клетки изменению отношения фосфолипидов к сфингомиелинам и липидов к белкам.
Избыток холестерина в мембранах, напротив, увеличивает микровязкость их бислоя фосфолипидных молекул, понижая скорость диффузии некоторых веществ через мембраны клеток.
Пища, обогащенная витаминами А, Е, С, Р улучшает обмен липидов в мембранах эритроцитов, снижает микровязкость мембран. Это повышает деформируемость эритроцитов, облегчает выполнение ими транспортной функции (глава 6).
Дефицит жирных кислот и холестерина в пище нарушает липидный состав и функции мембран клеток.
Например, дефицит жиров нарушает функции мембраны нейтрофилов, что угнетает их способность к движению и фагоцитозу (активный захват и поглощение микроскопических инородных живых объектов и твердых частиц одноклеточными организмами или некоторыми клетками).
В регулировании липидного состава мембран и их проницаемости, регуляции пролиферации клеток важную роль играют активные формы кислорода, образующиеся в клетке сопряженно с нормально протекающими метаболическими реакциями (микросомальным окислением и др.).
Образующиеся активные формы кислорода - супероксидный радикал (О 2), перекись водорода (H 2 О 2) и др. представляют собой чрезвычайно реакционноспособные вещества. Их основным субстратом в реакциях свободнорадикального окисления являются ненасыщенные жирные кислоты, входящие в состав фосфолипидов мембран клетки (так называемые реакции перекисного окисления липидов). Интенсификация этих реакций может вызвать повреждение мембраны клетки, ее барьерной, рецепторной и обменной функций, модификацию молекул нуклеиновых кислот и белков, что ведет к мутациям и инактивации ферментов.
В физиологических условиях интенсификация перекисного окисления липидов регулируется антиоксидазной системой клеток, представленной ферментами, инактивируюшими активные формы кислорода - супероксиддисмутазой, каталазой, пероксидазой и веществами, обладающими антиокислительной активностью - токоферолом (витамин Е), убихиноном и др. Выраженный защитный эффект на мембраны клетки (цитопротекторный эффект) при различных повреждающих воздействиях на организм оказывают простагландины Е и J2, «гася» активацию свободнорадикального окисления. Простагландины защищают слизистую желудка и гепатоциты от химических повреждений, нейроны, клетки нейроглии, кардиомиоциты - от гипоксических повреждений, скелетные мышцы — при тяжелой физической нагрузке. Простагландины, связываясь со специфическими рецепторами на клеточных мембранах стабилизируют бислой последних, уменьшают потерю мембранами фосфолипидов.
Функции рецепторов мембран
text_fields
text_fields
arrow_upward
Химический или механический сигнал вначале воспринимается рецепторами мембраны клетки. Следствием этого является химическая модификация мембранных белков, влекущая активацию «вторичных посредников», обеспечивающих быстрое распространение сигнала в клетке к ее геному, энзимам, сократительным элементам и т.д.
Схематично трансмембранная передача сигнала в клетке может быть представлена следующим образом:
1) Возбужденный воспринятым сигналом рецептор активирует у — белки мембраны клетки. Это происходит при связывании ими гуанозинтрифосфата (ГТФ).
2) Взаимодействие комплекса «ГТФ-у- белки», в свою очередь, активирует фермент - предшественник вторичных посредников, расположенный на внутренней стороне мембраны.
Предшественником одного вторичного посредника - цАМФ, образующегося из АТФ, является фермент аденилатциклаза;
Предшественником других вторичных посредников - инозитолтрифосфата и диацилглицерина, образующихся из фосфатидилинозитол-4,5-дифосфата мембраны, является фермент фосфолипаза С. Кроме того, инозитолтрифосфат мобилизует в клетке еще один вторичный посредник - ионы кальция, участвующие практически во всех регуляторных процессах в клетке. Так, например, образовавшийся инозитолтрифосфат вызывает выброс кальция из эндоплазматического ретикулума и повышение его концентрации в цитоплазме, тем самым включая различные формы клеточного ответа. С помощью инозитолтрифосфата и диацилглицерина регулируется функция гладких мышц и В-клеток поджелудочной железы ацетилхолином, передней доли гипофиза тиреогропин-релизинг фактором, ответ лимфоцитов на антиген и т.д.
В некоторых клетках роль вторичного посредника выполняет цГМФ, образующийся из ГТФ с помощью фермента гуанилатциклазы. Он служит, например, вторичным посредником для натрийуретического гормона в гладких мышцах стенок кровеносных сосудов. цАМФ служит вторичным посредником для многих гормонов - адреналина, эритропоэтина и др. (глава 3).
Клеточная мембрана
Изображение клеточной мембраны. Маленькие голубые и белые шарики соответствуют гидрофобным «головкам» фосфолипидов, а присоединённые к ним линии - гидрофильным «хвостам». На рисунке показаны только интегральные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны - молекулы холестерола Жёлто-зеленые цепочки бусинок на наружной стороне мембраны - цепочки олигосахаридов , формирующие гликокаликс
Биологическая мембрана включает и различные белки : интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов .
Функции
- барьерная - обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов . Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
- транспортная - через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке оптимального и концентрации ионов, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине неспособные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза .
При пассивном транспорте вещества пересекают липидный бислой без затрат энергии по градиенту концентрации путем диффузии. Вариантом этого механизма является облегчённая диффузия , при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза , которая активно вкачивает в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+). - матричная - обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие.
- механическая - обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки , а у животных - межклеточное вещество .
- энергетическая - при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
- рецепторная - некоторые белки, находящиеся в мембране, являются рецепторами (молекулами, при помощи которых клетка воспринимает те или иные сигналы).
Например, гормоны , циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней. - ферментативная - мембранные белки нередко являются ферментами . Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
- осуществление генерации и проведения биопотенциалов .
С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса . - маркировка клетки - на мембране есть антигены , действующие как маркеры - «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды , гликолипиды и холестерол . Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим - более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп ) затруднён.
Мембранные органеллы
Это замкнутые одиночные или связанные друг с другом участки цитоплазмы , отделённые от гиалоплазмы мембранами. К одномембранным органеллам относятся эндоплазматическая сеть , аппарат Гольджи , лизосомы , вакуоли , пероксисомы ; к двумембранным - ядро , митохондрии , пластиды . Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Избирательная проницаемость
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза , аминокислоты , жирные кислоты , глицерол и ионы , причем сами мембраны в известной мере активно регулируют этот процесс - одни вещества пропускают, а другие нет. Существует четыре основных механизма для поступления веществ в клетку или вывода их из клетки наружу: диффузия , осмос , активный транспорт и экзо- или эндоцитоз . Два первых процесса носят пассивный характер, то есть не требуют затрат энергии; два последних - активные процессы, связанные с потреблением энергии.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами - интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия .
См. также
Литература
- Антонов В. Ф., Смирнова Е. Н., Шевченко Е. В. Липидные мембраны при фазовых переходах. - М .: Наука, 1994.
- Геннис Р. Биомембраны. Молекулярная структура и функции: перевод с англ. = Biomembranes. Molecular structure and function (by Robert B. Gennis). - 1-е издание. - М .: Мир, 1997. - ISBN 5-03-002419-0
- Иванов В. Г., Берестовский Т. Н. Липидный бислой биологических мембран. - М .: Наука, 1982.
- Рубин А. Б. Биофизика, учебник в 2 тт . - 3-е издание, исправленное и дополненное. - М .: издательство Московского университета, 2004. -





